

Eukaryogenesis
Copyright © 2019-2020 C.E. by Dustin Jon Scott
The Domain Eukaryota / clade eucarya is...
Part I.b.
The Role of The Mitochondrion in Eukaryogenesis
Mitochondria are nigh-ubiquitous among the eukarya. Even microbial eukaryotes who were once believed to have diverged from the other eukarya before the acquisition of mitochondria have their own equivalent ATP- and ADP-synthesizing organelles that now appear to be derived from a mitochondrial ancestor, making it probable that the common ancestor of all known eukaryotes possessed a mitochondrion (Embley et al., 2003). Furthermore, the ATP-sensing kinase AMPK is found in all eukaryotic cells (Hardie et al. 2003).
"Along the lineage leading to LECA, what we might choose to designate as the first eukaryotic common ancestor (FECA) (11) depends (barring that miraculous cataclysm) on which eukaryote-defining feature we take as necessary and sufficient for eukaryoteness. Nowadays, many theorists would nominate the mitochondrion as that feature.
(Booth & Doolittle, 2015)
(Lopez-Garcia & Moreira, 2015)
Part I.b-1.)
Symbiogenesis: Endosymbiotic Theory
Part I.b-2.)
Other Theories
Part I.c.)
The Timing of Mitochondrial Acquisition in Eukaryogenesis
"The evolutionary origins of the oldest components of the AMPK pathway, however, extend into the pre-eukaryotic era, and descendants of these ancient proteins can still be found in contemporary prokaryotes. The TOR complex in turn appears as a eukaryotic invention, possibly to aid in retrograde signaling between the mitochondria and the remainder of the cell," (Roustan et al., 2016)"
Because the AMPK pathway has not been conserved among the prokaryotes, it may be inferred that in general it is highly variable and that such a dramatic anatomical restructuring as nucleus-cytosol compartmentilization would not likely have kept this system intact were it not constrained by the presence of mitochondria in the eukaryotic cell. This may be interpreted as indicating that the acquisition of the mitochondria either antedates or coincided with nucleus-cytosol compartmentalization.
"The central finding is that a subset of archaea encode actins that are not only monophyletic with eukaryotic actins but also share unique structural features with actin-related proteins (Arp) 2 and 3. All phagocytic processes are strictly dependent on remodeling of the actin cytoskeleton and the formation of branched filaments for which Arp2/3 are responsible. The presence of common structural features in Arp2/3 and the archaeal actins suggests that the common ancestors of the archaeal and eukaryotic actins were capable of forming branched filaments, like modern Arp2/3. The Rho family GTPases that are ubiquitous regulators of phagocytosis in eukaryotes appear to be of bacterial origin, so assuming that the host of the mitochondrial endosymbiont was an archaeon, the genes for these GTPases come via horizontal gene transfer from the endosymbiont or in an earlier event," (Yutin et al., 2009).
Part I.c-1.)
“Phagocytosis First” Hypothesis
"Phagocytosis, that is, engulfment of large particles by eukaryotic cells, is found in diverse organisms and is often thought to be central to the very origin of the eukaryotic cell, in particular, for the acquisition of bacterial endosymbionts including the ancestor of the mitochondrion," (Yutin et al., 2009).
Part I.c-1A.)
Archezoan Hypothesis
"Under the so-called archezoan hypothesis, the organism that acquired the endosymbiont was a proto-eukaryote (dubbed the archezoan) that already possessed the nucleus, the endomembrane system, the cytoskeleton, and other hallmark structures of the eukaryotic cell [5,7,8]. In other words, the hypothetical archezoan is envisaged as an amitochondrial, unicellular eukaryotic organism. The major difficulty faced by the archezoan hypothesis is that so far all candidate archezoa, such as Diplomonada, Parabasalia, and Microsporidia, have been shown to possess organelles derived from or, at least, related to mitochondria (hydrogenosomes, mitosomes, and others) as well as some nuclear genes of apparent mitochondrial (alpha-proteobacterial) origin [1,6]. Thus, the proponents of the archezoan hypothesis are forced to postulate that the archezoa represent an extinct lineage of primitive eukaryotes [8]," (Yutin et al., 2009).
Part I.c-1B.)
Phagocytosing Archaeon Theory (phAT)
Part I.c-2.)
“Symbiogenesis First” Hypothesis
States that the original host cell which acquired the mitochondrion was an archaeon.
“The hypotheses that oppose the archezoan concept are symbiotic scenarios in which the mitochondrial endosymbiosis is seen as the event that triggered eukaryogenesis in the first place. This idea traces back to the classic 1967 paper of Sagan (Margulis) [4] but received a major boost from the discovery of mitochondria-related organelles and genes of apparent mitochondrial origin in all thoroughly characterized eukaryotic cells [1,9,10]. Under the symbiotic scenarios that differ in details, the host that engulfed the alpha-proteobacterial ancestor of the mitochondria is posited to have been not a proto-eukaryote but rather an archaeon that closely resembled the currently known archaea, at least, in terms of the cell organization [1,11-13]. The advantage of the symbiotic scenarios is that they provide plausible, even if rather general explanations for the origin of the remarkable organizational and functional complexity of the eukaryotic cell as a result of diverse interactions between the host and the endosymbiont. However, the potentially serious difficulty faced by these scenarios is that prokaryotes have no known mechanisms for engulfing other prokaryotic cells (although at least one case of endosymbiosis among bacteria has been reported [14]). Thus, under these scenarios, the symbiosis between two prokaryotic cells would depend on an extremely rare, if not unique, spurious event � the "fateful encounter" hypothesis using the memorable phrase of De Duve [15]." (Yutin et al., 2009)
Part I.c-3.)
Ectosymbiotic Precursor?
Part II
Origin of The Mitochondrion
The origin of mitochondria is of vital importance to understanding eukaryotic evolution.
Arcaryan Hypothesis
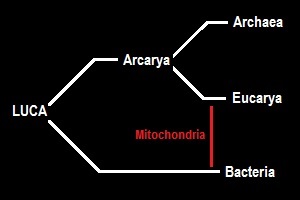
This hypothesis easily accomodates the Archaeozoan hypothesis as well as other “Phagocytosis first” scenarios, which postulate distinctively eukaryotic features in the host cell which acquired the mitochondrial ancestor.
TACK & The Eocyte Tree Fusion Scenario
Places the ancestor of the Eukaryotes within the Archaea rather than merely defining them as sister-groups.
TACK (Thaumarchaeota-Aigarchaeota-Crenarchaeota-Korarchaeota) superphylum (Guy & Ettema, 2011)
Asgardian Herritage
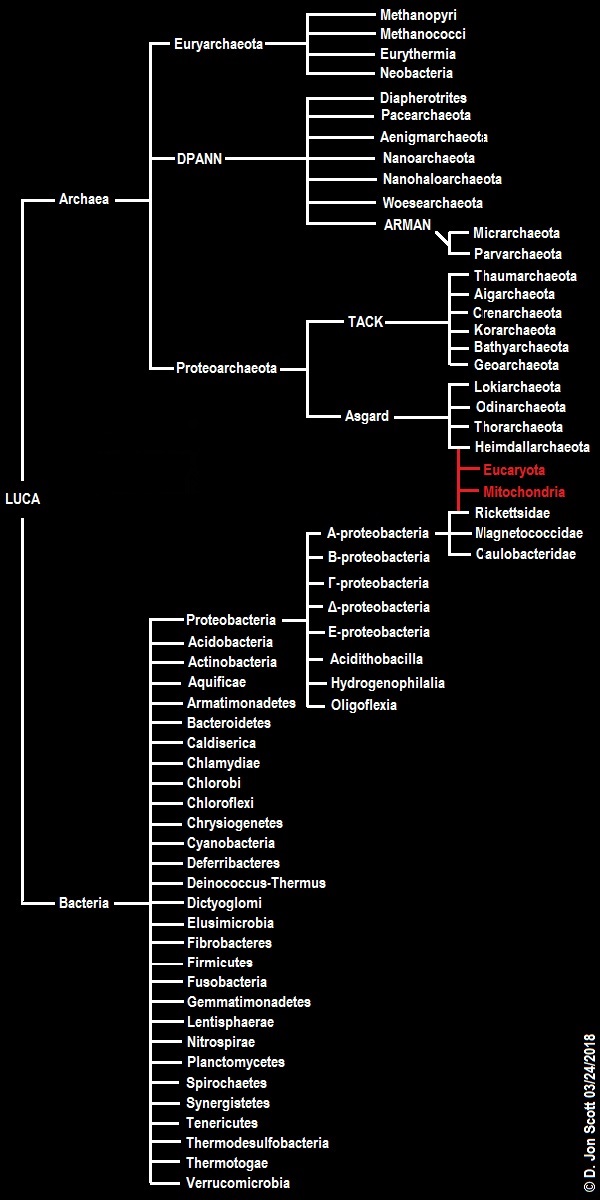
It is tempting to speculate that the archaeal ancestor may have been a multicellular archaeon capable of forming biofilms with some limited cellular differentiation, not wholey unlike Methanosarcina acetivorans, and that the acquisition of the mitochondrion permitted eukaryotic gene regulation and therefore complex multicellularity. However, since there are a large number of unicellular and facultatively multicellular eukaryotes, it's just as likely that facultative multicellularity in eukaryotes, bacteria, and archaea, is an example of convergent evolution.